L'analisi del doppio ibrido è una tecnica molecolare ampiamente utilizzata per studiare le interazioni proteina-proteina all'interno di una cellula. Queste interazioni sono fondamentali per quasi tutti i processi biologici e la loro comprensione è cruciale per svelare i meccanismi molecolari alla base della vita e delle malattie.

Un Breve Ripasso: Il Lievito Saccharomyces cerevisiae e il Metabolismo del Galattosio
Per comprendere appieno il funzionamento dell'analisi del doppio ibrido, in particolare nella sua applicazione più comune, il sistema a due ibridi di lievito (Y2H), è utile richiamare brevemente alcuni concetti sul metabolismo del galattosio in Saccharomyces cerevisiae. Questo lievito, infatti, utilizza il glucosio come fonte di energia principale. In presenza di glucosio, i geni coinvolti nel catabolismo del galattosio vengono repressi. Tuttavia, in condizioni di carenza di glucosio e abbondanza di galattosio, quest'ultimo entra nella cellula grazie a un recettore di membrana, il cui gene codificante è espresso costantemente a livello basale. Il galattosio funge quindi da induttore del sistema, attivando l'espressione dei geni necessari per il suo metabolismo.
Il Fattore di Trascrizione Gal4: Il Cuore del Sistema
Il fattore di trascrizione Gal4 è un elemento chiave nel meccanismo del doppio ibrido. Possiede due domini fondamentali per la sua funzione, localizzati in punti diversi della sua struttura proteica. La sua regione N-terminale include un dominio a dita di zinco, responsabile del legame al DNA, mentre la regione C-terminale contiene un dominio di trans-attivazione, essenziale per l'attivazione trascrizionale. La natura modulare di questi due domini è cruciale: il funzionamento del fattore di trascrizione è garantito anche dalla sola vicinanza spaziale dei domini, senza la necessità di un legame diretto tra loro. Questa caratteristica è stata abilmente sfruttata per sviluppare la tecnica del doppio ibrido.
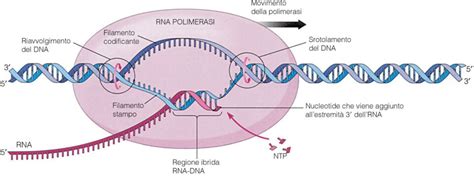
Come Funziona l'Analisi del Doppio Ibrido: Il Sistema Y2H
Inventato da Fields e Song nel 1989, lo screening in doppio ibrido in lievito costituisce tutt'oggi il metodo di screening più rapido, semplice e utilizzato per l'identificazione di interazioni proteina-proteina. Questo sistema si basa sulla ricombinazione della funzionalità del fattore di trascrizione Gal4.
Proteine "Bait" e "Prey"
Nel sistema Y2H, una delle proteine di interesse (chiamata spesso proteina "esca" o "bait") viene fusa con il dominio di legame al DNA (DB) di Gal4. L'altra proteina di interesse (chiamata "preda" o "prey") viene fusa con il dominio di trans-attivazione (AD) di Gal4. Queste due proteine di fusione vengono co-espresse in un ceppo di lievito che contiene uno o più geni reporter a valle di una sequenza di attivazione a monte GAL1.
Rilevazione dell'Interazione
Se le due proteine di interesse interagiscono tra loro, portano in stretta vicinanza il dominio di legame al DNA e il dominio di trans-attivazione. Questa associazione ricrea un fattore di trascrizione chimerico funzionale, che attiva l'espressione dei geni reporter. I reporter sono normalmente di due tipi: marcatori prototrofici, come HIS3 o ADE2, che consentono la sopravvivenza su terreno di crescita impoverito di specifiche sostanze nutritive, o enzimi cromogenici come la ß-galattosidasi, che possono essere utilizzati per la selezione tramite colorazione blu/bianca delle colonie. Se le proteine non interagiscono, i domini di Gal4 rimangono separati, il fattore di trascrizione non si ricompone e il gene reporter non viene espresso.
Screening in batch del sistema a due ibridi di lievito per confrontare le interazioni proteiche |...
Screening di Interazioni
Questa tecnica viene spesso utilizzata per condurre dei veri e propri screening, utilizzando diverse combinazioni di proteine bait e prey. Ad esempio, per identificare nuovi interattori di una proteina di interesse A, il cDNA del dominio di legame di Gal4 viene fuso al cDNA della proteina A, e il cDNA del dominio attivatore di Gal4 viene fuso a una libreria di cDNA (B) contenente potenziali proteine interattrici.
Vantaggi e Limitazioni dell'Y2H
Il test del doppio ibrido di lievito è facile da eseguire. Tuttavia, presenta una limitazione significativa: può essere utilizzato solo per studiare le interazioni tra proteine con localizzazione nucleare.
Variazioni sul Tema: Altri Metodi per Studiare le Interazioni Proteina-Proteina
Il sistema a doppio ibrido di lievito è solo uno dei molti metodi in vivo sviluppati per studiare le interazioni proteina-proteina. Questi metodi possono essere classificati in base alle proprietà del sistema reporter utilizzato. Tutti si basano su un sistema reporter costituito da due parti: una collegata a una delle proteine di interesse (esca) e l'altra all'altra proteina di interesse (preda). Quando le proteine interagiscono, le due parti del sistema reporter vengono portate in stretta vicinanza, producendo un effetto chiaramente rilevabile.
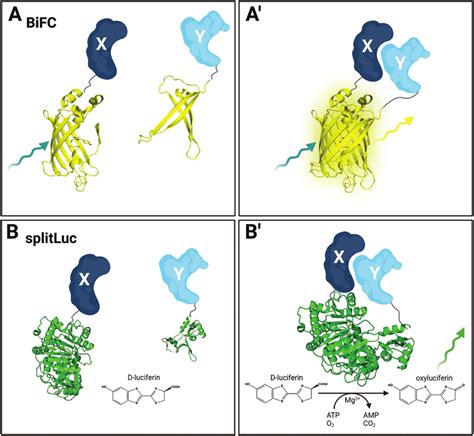
Assays di Complementazione dei Frammenti Proteici (PCA)
Nei PCA, il reporter è una singola proteina che è stata scissa in due frammenti. I frammenti, di per sé, non sono funzionali. Tuttavia, la proteina può ricostituirsi quando entrambi i frammenti vengono portati in stretta vicinanza, ripristinando la sua funzionalità.
Test di Complementazione della Luciferasi Divisa
Ad esempio, nel test di complementazione della luciferasi divisa, la molecola della luciferasi viene divisa in due frammenti. Nessuno dei due frammenti può produrre luce autonomamente, ma se sono sufficientemente vicini, la luciferasi viene ricostituita e può nuovamente emettere luce. Questo metodo è molto sensibile e offre un elevato rapporto segnale-rumore, poiché la maggior parte degli organismi modello non è luminescente. Sia la luciferasi di Firefly che la luciferasi di Renilla sono state utilizzate con successo per studiare le interazioni proteina-proteina in vivo. A differenza di altri PCA, la ricostituzione della luciferasi è reversibile nella maggior parte dei casi, rendendo il metodo adatto per monitorare le interazioni dinamiche in tempo reale. L'implementazione della luciferasi divisa è un metodo popolare per studiare le interazioni proteina-proteina nelle piante, utilizzando un sistema di imaging vegetale in vivo per visualizzare la luminescenza sulle foglie trasformate.
BiFC (Bimolecular Fluorescence Complementation)
BiFC è un PCA che utilizza una proteina fluorescente scissa (come GFP o altre). I frammenti della proteina non sono fluorescenti, ma la proteina fluorescente viene ricostituita quando entrambi i frammenti vengono portati in stretta vicinanza. Simile a FRET, BiFC è un metodo fluorescente per misurare le interazioni proteina-proteina, ma presenta una differenza importante: la complementazione dei frammenti proteici fluorescenti tagliati è irreversibile, rendendo il metodo inadatto per indagare le dinamiche di interazione. Un'altra complicazione è che i frammenti scissi alla fine si ricompongono da soli, il che aumenta la rilevazione di falsi positivi e richiede estrema cura nella pianificazione dell'esperimento e nella progettazione di controlli appropriati.
Sistemi Basati sull'Espressione Genica
In questi metodi, se le due proteine di interesse interagiscono, la combinazione del dominio di legame al DNA e del dominio di transattivazione guida l'espressione del reporter. Oltre al test del doppio ibrido di lievito, il sistema split-ubiquitin è un altro esempio basato sull'espressione genica.
Sistema Split-Ubiquitin
Questo sistema è concettualmente simile al sistema a doppio ibrido di lievito ed è altrettanto facile da implementare. Tuttavia, ha l'ulteriore vantaggio di poter essere applicato praticamente a qualsiasi proteina, non solo a quelle nucleari. Il sistema split-ubiquitin è una specie di PCA: la proteina ubiquitina viene divisa in due frammenti non funzionali, ognuno dei quali è fuso con un membro della coppia di proteine di interesse. Quando i due frammenti interagiscono, l'ubiquitina si riassembla e diventa funzionale. Questo promuove la scissione tramite la proteasi ubiquitina-specifica, portando all'attivazione di un gene reporter. Tuttavia, i reporter basati sull'espressione genica non sono in grado di seguire i cambiamenti dinamici nell'interazione e possono essere quantificati solo mediante assorbimento e utilizzando un enzima cromogenico. Possono essere utilizzati per valutare un gran numero di interazioni binarie mescolando due tipi di accoppiamento aploidi che sono stati trasformati con diversi set di proteine di interesse.
Metodi Basati sulla Fluorescenza e Bioluminescenza
Le proteine fluorescenti e i sistemi luminescenti sono ampiamente utilizzati per rilevare le interazioni proteina-proteina.
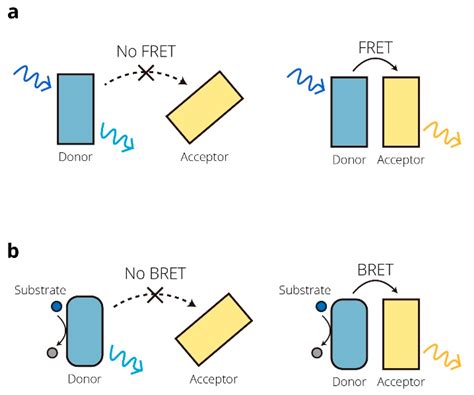
FRET (Förster Resonance Energy Transfer)
Il FRET è un metodo di fluorescenza ampiamente utilizzato. In questo approccio, una proteina donatrice fluorescente è accoppiata a una delle proteine di interesse e una proteina accettore all'altra proteina. Se non c'è interazione, dovrebbe essere rilevata solo la fluorescenza del donatore. Tuttavia, se si verifica un'interazione e le due proteine fluorescenti si trovano a una distanza di 10 nm o inferiore, la proteina donatrice fluorescente può eccitare l'accettore tramite trasferimento di energia di risonanza (accoppiamento dipolo-dipolo). È normale che una certa emissione dell'accettore venga rilevata anche senza interazione, a causa di una sovrapposizione spettrale nell'eccitazione delle due proteine fluorescenti. Per questo, il rapporto FRET (emissione dell'accettore divisa per l'emissione del donatore) viene normalmente utilizzato per la quantificazione, con un aumento significativo del rapporto che indica un'interazione. Questo tipo di FRET è talvolta indicato come FRET a emissione sensibilizzata (SE-FRET o seFRET). Poiché SE-FRET è il metodo comune per misurare FRET con un lettore di micropiastre, di solito viene indicato semplicemente come FRET. Altri tipi di FRET utilizzati con i microscopi a fluorescenza includono FLIM FRET e il photobleaching dell'accettore FRET (dove l'aumento di fluorescenza del donatore viene misurato dopo il fotobleaching dell'accettore). Come la maggior parte dei metodi basati sull'intensità della fluorescenza, FRET soffre di un alto fondo e quindi di bassi rapporti segnale-rumore, che ne limitano la sensibilità.
TR-FRET (Time-Resolved Förster Resonance Energy Transfer)
Per superare le limitazioni di FRET, sono stati sviluppati metodi alternativi come TR-FRET. In TR-FRET, i fluorofori con una vita di fluorescenza molto lunga (principalmente chelati di lantanidi, come europio, terbio e samario) vengono utilizzati per bypassare l'interferenza di molecole con una breve durata di fluorescenza o altri fattori (in particolare l'eccitazione della luce). Invece di misurare la luce quando la luce di eccitazione è accesa, le misure TR-FRET vengono eseguite centinaia di microsecondi dopo. I chelati possono ancora emettere luce, ma tutti gli altri fluorofori nel campione e ovviamente la luce di eccitazione sono già svaniti. La fluorescenza risolta nel tempo (TRF) non deve essere confusa con la durata della fluorescenza.
FLIM FRET (Fluorescence Lifetime Imaging Microscopy FRET)
La durata della fluorescenza descrive il tempo medio che una proteina fluorescente trascorre nello stato eccitato prima di tornare allo stato fondamentale emettendo un fotone. I metodi basati sulla durata della fluorescenza si basano sulla misura della durata dei singoli fluorofori piuttosto che sui loro spettri di emissione. Poiché la durata della fluorescenza non dipende dalla concentrazione, dall'assorbimento del campione, dall'intensità di eccitazione o dall'emorragia spettrale, è più robusta dei metodi basati sull'intensità. Un fenomeno interessante è che il FRET non solo aumenta l'emissione di fluorescenza dell'accettore, ma riduce anche la durata della fluorescenza del donatore a causa del trasferimento di energia. La combinazione con FLIM per visualizzare FRET consente di studiare le interazioni proteina-proteina in vivo con elevata affidabilità, consentendo una risoluzione spazio-temporale altamente dinamica. Tuttavia, un uso corretto di FLIM richiede componenti aggiuntivi hardware sofisticati e una formazione approfondita. Le misure del tempo di vita della fluorescenza possono anche essere eseguite con un fluorimetro.
BRET (Bioluminescence Resonance Energy Transfer)
BRET è stato sviluppato per bypassare alcuni dei problemi di fondo di FRET. Invece di utilizzare una proteina fluorescente come donatore, viene utilizzata una luciferasi. Poiché le luciferasi producono luce attraverso una reazione chimica, non è necessaria una sorgente di luce esterna per l'eccitazione. BRET ha quindi un fondo molto piccolo e non soffre di problemi spesso associati al FRET, come autofluorescenza, dispersione della luce o sbiancamento fotografico. La classificazione di BRET come metodo di luminescenza è quindi controversa: mentre il segnale misurato rappresenta l'emissione di una proteina fluorescente (es. eYFP per BRET1 e GFP per BRET2), la lampada di eccitazione non può essere utilizzata (poiché l'accettore è eccitato dalla luciferasi del donatore tramite accoppiamento dipolo-dipolo).
Analisi del Doppio Ibrido: Non Solo Interazioni Proteiche
È importante notare che l'espressione "analisi del doppio ibrido" può riferirsi anche al "duo test" in ambito prenatale. Sebbene il nome sia simile, le due tecniche sono completamente diverse.
Il Duo Test in Ostetricia: Uno Screening Preliminare
Il duo test è un'analisi di screening utilizzata nella popolazione ostetrica per individuare le gravidanze a rischio di trisomia 21 (sindrome di Down) o altre anomalie cromosomiche, come la trisomia 18. Non ha finalità diagnostiche, ma permette di individuare le gestanti con un "alto" rischio di portare in grembo un feto interessato da queste condizioni. L'aggettivo "alto" è posto tra virgolette perché, nella maggior parte dei casi, le probabilità che il feto sia malato sono esigue, ma comunque giustificano ulteriori approfondimenti diagnostici. La parola "screening" significa "scegliere attentamente".
Componenti del Duo Test
Il duo test è un'analisi ematica che misura i livelli di due ormoni prodotti dalla placenta materna: la beta-hCG libera e la PAPP-A (Pregnancy Associated Plasma Protein-A). Nelle gravidanze con feto affetto da sindrome di Down, la beta-hCG libera risulta elevata rispetto alle gravidanze con feto euploide (non affetto da sindrome di Down), mentre la PAPP-A risulta inferiore alla norma.
Scopo e Limitazioni
Lo scopo del duo test non è diagnosticare la malattia, bensì identificare le gravidanze a maggior rischio in modo da eseguire ulteriori indagini a fini diagnostici. Per confermare il sospetto di anomalia cromosomica è necessario sottoporsi ad esami invasivi, come la villocentesi o l'amniocentesi. La ragione per cui le gestanti non vengono direttamente sottoposte a questi esami, evitando le analisi preliminari di screening come il duo test, è legata al rischio intrinseco di abortività che accompagna queste procedure invasive. Le probabilità di perdere il feto a seguito di queste manovre diagnostiche, infatti, sono contenute, ma non trascurabili (0,5-1%).
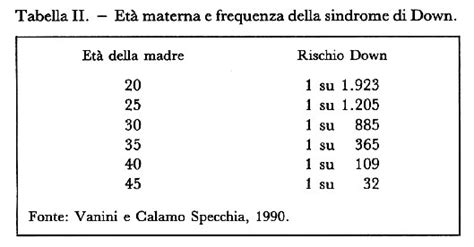
Età Materna e Rischio
Il rischio di aborto legato alla villocentesi o all'amniocentesi è considerato accettabile per le donne con più di 35 anni, ma non per le gestanti più giovani. Il rischio di dare alla luce un figlio affetto da sindrome di Down aumenta all'aumentare dell'età materna. Se è vero che le donne con più di 35 anni sono esposte al maggior rischio di avere figli affetti da sindrome di Down, è altrettanto vero che la maggior parte (circa il 70%) dei bambini con trisomia 21 nasce da donne più "giovani", considerate a basso rischio. Questa apparente contraddizione si basa sull'evidenza statistica che la maggior parte delle gravidanze interessa donne di età inferiore a 35 anni; di conseguenza, facendo molti più figli, in termini assoluti le fasce più giovani della popolazione danno alla luce un maggior numero di bambini affetti da mongolismo e altre anomalie cromosomiche.
Indicazioni per il Duo Test
Il duo test è raccomandato per le donne più giovani per identificare le gravidanze a rischio prima della nascita. Non è di regola indicato nelle donne con età superiore a 35 anni, per le quali l'età stessa è già sufficiente a classificarle come "ad alto rischio", rendendo giustificati altri accertamenti invasivi. Altri fattori che influenzano il rischio includono l'abitudine al fumo, la tendenza alla minaccia d'aborto e precedenti figli con anomalie cromosomiche.
Interpretazione dei Risultati
I risultati del duo test vengono espressi in percentuali (ad esempio 1 caso patologico possibile su 1000 o un caso patologico possibile su 50). Se il numero è compreso tra 1/1 e 1/350, la probabilità che il bambino sia affetto da Trisomia 21 è considerata elevata. Nonostante l'ultimo estremo (una probabilità su 350) mostri un rischio tutto sommato contenuto, lo si considera comunque appropriato per ulteriori approfondimenti.
Efficacia del Duo Test
Nonostante la sua utilità, il duo test è lontano dall'essere un esame di screening perfetto. La sua esecuzione permette di individuare soltanto l'ottanta percento dei feti interessati da sindrome di Down. Ciò significa che tra tutte le gravidanze complicate da trisomia ventuno, solamente l'80% viene considerata a rischio e, di conseguenza, inviata ad amniocentesi o villocentesi. Fortunatamente, l'amniocentesi o la villocentesi sono in grado di riconoscere la presenza di anomalie praticamente nella totalità (>99%) dei feti esaminati.
Test Combinato e Quad Test
Un ulteriore aumento della sensibilità si ottiene affiancando il duo test a un esame ecografico chiamato translucenza nucale; in questo caso si parla di test combinato. Un ulteriore aumento della sensibilità (fino al 95%) si ottiene affiancando duo test, translucenza nucale e quad test.