Le Lamiaceae Martinov, 1820, una famiglia di piante spermatofite dicotiledoni, si distinguono per i loro fiori caratteristici a forma bilabiata, appartenenti all'ordine delle Lamiales. Questo vasto gruppo botanico trova il suo epicentro di diversificazione nel bacino del Mediterraneo, prosperando in ambienti degradati come la macchia mediterranea e le garighe, su substrati rocciosi, calcarei o sabbiosi. Il nome stesso della famiglia deriva dal suo "genere tipo", Lamium (tribù Lamieae). Già Gaio Plinio Secondo, illustre naturalista latino, suggerì un'etimologia legata al greco "laimos", che significa "fauci - gola", evocando la forma distintiva del fiore. Altre possibili origini greche includono "lamos" (larga cavità) o persino il nome di una regina libica, Làmina.
Il nome scientifico della famiglia fu inizialmente definito "Labiate" dal botanico francese Antoine Laurent de Jussieu nel 1789, per poi essere perfezionato nella forma attuale (Lamiaceae) dal botanico russo Ivan Ivanovič Martinov.
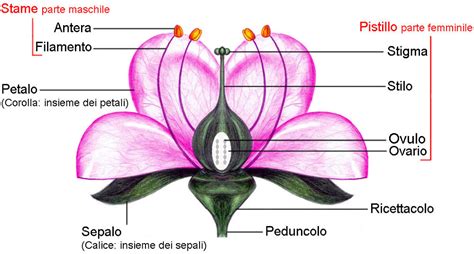
Il portamento delle specie Lamiaceae è prevalentemente erbaceo, annuale o perenne. Le forme biologiche più comuni sono emicriptofite e terofite, ma non mancano specie camefite, nanofanerofite, cespugliose e, in alcuni casi, arboree, arbustive o lianose, con una generale tendenza xerofita. L'indumento è tipicamente pubescente, formato da peli semplici. Il fusto, nella maggior parte delle specie, presenta una sezione quadrangolare, dovuta alla presenza di fasci di collenchima nei quattro vertici, mentre le quattro facce sono concave. Raramente, nelle ascelle fogliari, si osservano spine. Le foglie sono disposte in modo opposto lungo il fusto, talvolta verticillate e decussate, e possono essere sessili o picciolate. La loro forma è variabile, da lanceolata a ovata, con margini interi, lobati o dentati.
Le infiorescenze sono di tipo tirsoide o racemoso, composte da verticilli ascellari sovrapposti. Ogni verticillo contiene da 2 a 20 fiori, disposti circolarmente e poggianti su due grandi brattee fogliose, che si staccano leggermente dall'infiorescenza vera e propria. Le brattee dei verticilli successivi sono disposte alternativamente. In alcune specie, le brattee sono subulate o spinose, e talvolta presentano colorazioni vivaci.
La Struttura Florale: Calice e Corolla Bilabiata
La formula fiorale delle Lamiaceae rivela adattamenti specifici per l'impollinazione. Il calice, gamosepalo e generalmente attinomorfo (a volte debolmente zigomorfo), è solitamente composto da 5 lobi uguali o subuguali, diritti e lanceolati o triangolari e spinescenti. In alcuni casi, il calice può essere debolmente bilabiato, con i lobi raggruppati in configurazioni come 2/3, 3/2, o 1/4.
La corolla, gamopetala e zigomorfa, assume la forma di un tubo dilatato distalmente, terminante in due evidenti labbra. La struttura tipica è 2/3, con un labbro posteriore (superiore) corto o moderatamente allungato, a forma di cappuccio, intero o smarginato al bordo, talvolta mucronato. Il labello (labbro inferiore) è piatto, patente, e generalmente formato da tre lobi, con quello centrale più grande, o talvolta tutti e tre di dimensioni uguali.
Esistono tuttavia eccezioni significative: nei generi Mentha, Lycopus e Ocimum, la corolla appare attinomorfa a 4 lobi. Nei generi Ajuga e Teucrium, il labbro superiore è assente. La corolla può presentarsi densamente pubescente. Le fauci interne sono spesso circondate da un anello di peli, un adattamento comune nelle "labiate" che funge da barriera per impedire l'accesso a insetti troppo piccoli e non adatti all'impollinazione.

Androceo e Gineceo: Organi Riproduttivi
L'androceo delle Lamiaceae è caratterizzato dalla presenza di quattro stami didinami, tutti fertili (raramente quelli posteriori sono sterili), inclusi o sporgenti dalle fauci della corolla e posizionati sotto il labbro superiore. Nei generi Lycopus, Salvia, Rosmarinus e Ziziphora, gli stami sono ridotti a due. I filamenti sono adnati alla corolla. Le antere sono ravvicinate a coppie, glabre o pubescenti, con due teche (biloculari) poco o ben distinte, parallele o divergenti. La deiscenza è longitudinale, raramente a valve. I granuli pollinici sono tipicamente da tricolpati a esacolpati.
Il gineceo presenta un ovario supero, profondamente 4-lobato, formato da due carpelli fusi (bicarpellare) e 4-loculare a causa della presenza di falsi setti divisori. La placentazione è assile. Sono presenti 4 ovuli (uno per presunto loculo), con un tegumento e tenuinucellati. Lo stilo, inserito alla base dell'ovario (ginobasico) o capitato, è filiforme. Il frutto è uno schizocarpo, composto da 4 nucule (o due per aborto) secche o carnose, arrotondate o alate all'apice, talvolta piatte e/o troncate. La superficie delle nucule può essere liscia, glabra, pubescente o tubercolata.
L'Impollinazione: Un Processo Fondamentale
La riproduzione nelle piante si basa sulla conservazione della specie attraverso la generazione di nuovi individui. La riproduzione sessuata, in particolare, permette il rimescolamento dei caratteri ereditari dei genitori, favorendo una maggiore adattabilità all'ambiente. Il processo cruciale che consente il trasferimento del polline dagli stami al pistillo è l'impollinazione.
L'impollinazione può essere di tipo autogamo (autoimpollinazione o autogamia), quando il polline feconda lo stigma dello stesso fiore o di fiori dello stesso individuo, o allogamo (impollinazione incrociata), quando il polline viene trasportato da un fiore all'organo femminile di un individuo diverso della stessa specie. Le piante che praticano l'autoimpollinazione sono dette autogame, mentre quelle che praticano l'impollinazione incrociata sono dette allogame.
Per evitare l'autoimpollinazione, le piante hanno sviluppato diversi meccanismi:
- Dioicia: Separazione dei sessi, con piante esclusivamente maschili e femminili.
- Dicogamia: Mancata contemporanea maturazione degli organi sessuati (protandria o protoginia).
- Ergogamia: Presenza di conformazioni organiche che impediscono al polline di raggiungere lo stigma dello stesso fiore.
- Eterostilia: La stessa specie presenta fiori con diversa lunghezza degli stili e degli stigmi.
- Autoincompatibilità (o Autosterilità): L'impossibilità genetica di produrre zigoti con il proprio polline, controllata da geni specifici (geni S).
LE SUPERSINTESI DI SCIENZE: le piante Angiosperme - il fiore e la fecondazione (classe prima)
Agenti Impollinatori: Vettori di Vita
Il trasporto del polline può avvenire tramite agenti abiotici (vento, acqua) o biotici (animali, principalmente insetti).
Impollinazione Abiotica
- Anemogama (o Anemofila): L'impollinazione avviene tramite il vento. Le piante anemofile producono grandi quantità di polline, generalmente liscio, di piccole dimensioni e polverulento, per aumentare le probabilità di successo. Gli stami sono spesso mobili, permettendo alle antere di disperdere il polline nell'aria. Gli stigmi sono piumosi per intercettare meglio il polline. Esempi includono molte graminacee e specie arboree con fiori raccolti in amenti.

- Idrogama (o Idrofila): L'impollinazione è facilitata dall'acqua. Questo tipo di impollinazione è meno diffuso e si osserva in alcune piante acquatiche come Ceratophyllum o Vallisneria. In quest'ultima, i fiori maschili si staccano e vengono trasportati dalle correnti ai fiori femminili. Il polline ha una germinazione difficile in ambiente artificiale per garantire la specificità.
Impollinazione Biotica (Zoogama)
L'impollinazione zoogama rappresenta una strategia evolutiva più recente e altamente specializzata, caratterizzata da una coevoluzione tra piante e animali impollinatori.
Entomogama (o Entomofila): È l'impollinazione effettuata dagli insetti. Le piante attirano gli insetti attraverso stimolanti ottici (pigmentazione e forma del perianzio) e chimici (odori emanati dai fiori). La fedeltà degli insetti a una specifica specie floreale è favorita dalla memorizzazione di queste sensazioni. Le caratteristiche dei fiori che favoriscono l'impollinazione entomogama includono:
- Polline: Più grande rispetto a quello anemofilo, con ornamentazioni superficiali e uno strato viscoso che ne facilita l'adesione alle setole degli insetti.
- Colori e Odori: Colori vivaci (blu, gialli, a volte con motivi ultravioletti invisibili all'uomo) e profumi specifici attraggono determinati insetti.
- Nettare: Secreto dai nettari, offre una ricompensa energetica agli impollinatori. La sua posizione e quantità influenzano quali insetti possono accedervi.
- Forma della Corolla: Varia da fiori "a disco" aperti, frequentati da molti impollinatori, a fiori "a tubo" o "chiusi", accessibili solo a insetti con organi boccali specifici (es. api con lingua lunga).
La coevoluzione ha portato a strategie sorprendenti:
- Mimetismo: Le orchidee del genere Ophrys imitano la forma e l'odore delle femmine di specifici insetti, ingannando i maschi che tentano l'accoppiamento e finendo per impollinare il fiore.
- Meccanismi a Trappola: Alcuni fiori, come quelli di Arum maculatum, intrappolano temporaneamente gli insetti per garantire l'impollinazione.
- Fiori per specifici impollinatori:
- Melittofili (api): Fiori con parti adatte all'appoggio, spesso blu o gialli, con segni ultravioletti. La salvia ne è un esempio, dove la corolla funge da "pista d'atterraggio".
- Psicofili (farfalle diurne): Fiori con colori vivaci e profumi dolci.
- Falenofili (farfalle notturne): Fiori bianchi o pallidi, che si aprono verso sera e emettono profumi forti e dolci dopo il tramonto.
- Cantofili (coleotteri): Fiori facilmente accessibili, con ovari inferi.
- Fiori visitati da mosche: A volte attratte da odori sgradevoli.

Ornitogama (o Ornitofila): L'impollinazione è effettuata dagli uccelli. I fiori ornitofili sono spesso vistosi (gialli o rossi), privi di zone di appoggio e producono abbondante nettare. Colibrì, nettarinie e alcuni pappagalli sono tra gli impollinatori più comuni.
Chirotterogama (o Chirotterofila): L'impollinazione è svolta dai pipistrelli. I fiori chirotterofili sono tipicamente grandi, robusti, si aprono di notte, sono bianchi o verdastri ed emanano odori forti, talvolta sgradevoli. Il baobab è un esempio di pianta impollinata da pipistrelli.
Altre forme di zoogamia: Sono stati documentati casi di impollinazione da parte di rettili (lucertole, gechi) e, in misura minore, molluschi.
Lamiaceae nell'Ecosistema dell'Impollinazione
La famiglia delle Lamiaceae, con la sua caratteristica corolla bilabiata, è perfettamente inserita nel complesso mondo dell'impollinazione entomogama. La forma bilabiata del fiore crea una sorta di "bocca" o "trappola" che guida l'insetto impollinatore verso il nettare e, al contempo, assicura il contatto con le antere e lo stigma. L'anello di peli nelle fauci, menzionato in precedenza, funge da ulteriore filtro, permettendo l'accesso solo agli insetti di dimensioni adeguate.
Molte Lamiaceae, come la salvia, presentano la corolla strutturata in modo da agire come una "pista d'atterraggio", facilitando l'avvicinamento dell'insetto. Durante la visita per raggiungere il nettare, l'insetto inevitabilmente si sporca di polline. La dimensione e la forma dei fiori delle Lamiaceae sono spesso ottimizzate per attrarre specifici gruppi di insetti, dimostrando un elevato grado di specializzazione in molti casi.
La diversità all'interno della famiglia Lamiaceae si riflette anche nelle strategie impollinative, con alcune specie che presentano adattamenti più marcati di altre. La comprensione di questi intricati meccanismi è fondamentale non solo per la biologia vegetale, ma anche per le applicazioni agricole, dove gli impollinatori svolgono un ruolo insostituibile nella produzione di cibo. La salute delle popolazioni di impollinatori è quindi intrinsecamente legata alla nostra sicurezza alimentare e alla biodiversità del pianeta.
tags: #impollinazione #entomofila #lamiaceae #schema #corolla #bilabiata